ウマの進化
当項目ではウマの進化の歴史について記述する。

現代の馬までの進化の軌跡は他の動物のものよりも化石の出土数も多く信頼性が高い。ウマ科を含む奇蹄目はK-Pg境界の後1000万年までの暁新世後期に誕生した。奇蹄目は元々、熱帯林での生活に順応していたが、バク科とサイ科が森に適応したのに対し、ウマは草原などステップ地帯での生活に適応した。その過程において、次第に背が高くなり、足指では中指の発達と並行して他の指の退化が進むなど、一定方向への系統的な変化が見て取りやすいことから、系統化石の好例とされる。
タイムライン
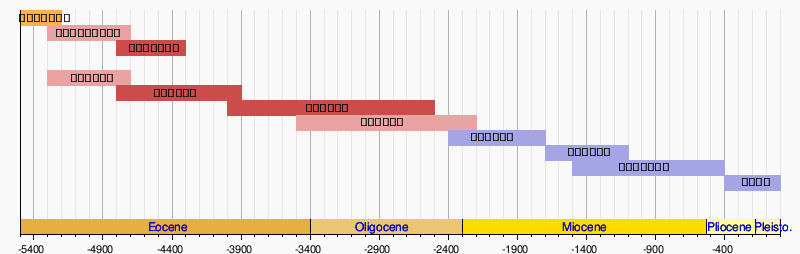
進化の過程
ヒラコテリウム

現在、最も古いと考えられているウマ科動物はヒラコテリウム Hyracotherium である。ヒラコテリウムの化石は18世紀にヨーロッパで見つかり、リチャード・オーウェンによって「ハイラックス(Hyrax)様の獣」を意味するヒラコテリウムと名づけられた。ヒラコテリウムはオスニエル・チャールズ・マーシュにより名づけられた「始新世のウマ」を意味するエオヒップス(Eohippus)という名も広まっている。ただし、正式な学名は優先順位の高い「ヒラコテリウム」となっている。
ヒラコテリウムは約5200万年前にはすでに北アメリカ大陸で生活していたとされている。体はキツネと同じくらいのサイズ(体高25~45センチメートル)で、比較的短く弾力性のある頭頸部とアーチ状の背骨を持っていた。歯は各側に切歯を3個、犬歯を1個、小臼歯を4個、大臼歯を3個備えており、合計で44個の歯を持っていた。大臼歯は葉を削りやすい形であり、ヒラコテリウムは葉食性(柔らかい木の葉や果物などを食べていた)である事が窺える。またヒラコテリウムは小さい脳を持っており、小さい前頭葉もあった。
すでに走ることに対しての進化は始まっており、手足は現在の馬のように体に比例して長かった。しかし、下肢骨のいくつかは不安定で、柔軟性に欠けていた。脚はそれぞれ5本ずつ指があったが、進化の過程で前肢は第1指が退化し4本、後肢は第1指と第5指が退化し3本になっている。爪先は犬のような鉤爪ではなく、小さなひづめがついていた。
約200万年の間に、ヒラコテリウムは進化し繁栄した。最も重要な進化がより葉食性に特化した歯の獲得である。始新世の間、ヒラコテリウムはウマ科の様々な種に分岐した。これらの完全な化石は北米(ワイオミング州ウィンド川など)で数多く発見された。また、現代のウマの先祖とは考えられていないパレオテリウム Propalaeotherium などの化石がヨーロッパでも見つかっている。プロパレオテリウムはパレオテリウム Palaeotherium へと進化するが、その後絶滅した。
オロヒップス

約5000万年前、始新世中期にヒラコテリウムはオロヒップス(Orohippus)へと進化した。オロヒップスとは山のウマを意味するが、実際にはオロヒップスは山には住んでいなかった。また、オロヒップスはプロトロヒップス(Protorohippus)という別名がある。体はヒラコテリウムと同じサイズだったが、より細い胴体、細長い頭、細い前肢、長い後足を持っていた。その体は跳躍力に優れていたと考えられている。
ヒラコテリウムとオロヒップスを分ける大きな変化は歯にあり、第一小臼歯が小さくなり、第三小臼歯は形を変えて大臼歯となった。また、歯冠はより大きくなり、より硬い植物もすり潰し、食べられるように進化した。
エピヒップス
約4700万年前、オロヒップスはより大きな臼歯を持つエピヒップス(Epihippus)へと進化した。デュシェーヌヒップス中間型(Duchesnehippus intermedius)と呼ばれた後期のエピヒップスには、漸新世のウマ科と同じ歯があった。
メソヒップス

始新世後期から漸新世(3200~2400万年前)初期に北米は乾燥するようになり、植物では初期のイネ科植物が出現し、森はイネ科などの草原へと変化してきた。砂で覆われていた平野に、現在のプレーリーに似た草原ができていった。
約4000万年前、始新世後期に環境の変化に適合し選択されたメソヒップス(Mesohippus)へと進化した。メソヒップスの体はエピヒップスより大きくなり、脚がより長くなった。隠れる場所の少ない平原では捕食者から逃げるためにより速く走る必要が出てきたからである。
漸新世初期にはメソヒップスは北米の広い範囲で生活していた。その脚は前後とも指が3本になっており、3つの爪先で歩行していた。前脚の第5指は退化し、第3指がより発達した。長く、細い脚から、メソヒップスは敏捷性に優れた動物であったことが窺える。
メソヒップスはエピヒップスと比べて、肩まで61センチメートルと少し大きく、また、背中はアーチ状ではなくなり、顔や鼻、首が長くなった。大きな大脳を持ち、頭蓋骨には現在の馬にも見られる小さな浅い窪みがある。その窪みは化石から馬の種の鑑定に使われている。メソヒップスは小臼歯が前部にあり、後のウマ科の動物が持つ6個の臼歯を持っていた。植物を食べるために、エピヒップスよりも硬く鋭い歯を獲得していた。
ミオヒップス
約3600万年前、メソヒップスへの進化のすぐ後にミオヒップス(Miohippus)は誕生している。メソヒップスとミオヒップスの間の化石はいくつか見つかっているが、ミオヒップスの登場は比較的突然であった。ミオヒップスはメソヒップスが段階的に進化したとされていたが、その後分岐進化であることが研究結果から明らかとなっている。今日ではミオヒップスはメソヒップスの亜種から進化し、長い間メソヒップスとミオヒップスが共存していたことが知られている。
ミオヒップスの体は大きく、脚の繋は少し変化した。顔の窪みはより大きく、より深くなった。上側臼歯はより硬い植物をすり潰すために動く歯冠を持った。
メソヒップスは漸新世中期に絶滅したのに対し、ミオヒップスは繁栄し中新世初期(2400~530万年前)に急速に様々な種に分化し始めた。分化した種は草原に適応した種と森林に適応した種の大きく2種類に分けられる。
カロバティップス

森に適応した種はカロバティップス(Kalobatippus)と呼ばれる種で、ミオヒップス中間型(Miohippus intermedius)とも呼ばれた。森では地面が柔らかいため、第2指と第4指は歩行に役立っており、再び発達していた。カロバティップスはアンキテリウム(Anchitherium)とも呼ばれた。カロバティップスはベーリング地峡を渡り、ヨーロッパやアジアまで進出した。カロバティップスはユーラシアでシノヒップス(Sinohippus)に、北米でハイポヒップス(Hypohippus)やメガヒップス(Megahippus)に進化した。ハイポヒップスは鮮新世初期に絶滅している。
パラヒップス
草原に残ったミオヒップスはパラヒップス(Parahippus)へ進化した。パラヒップスは小さいポニーほどの大きさで、現在の馬に似た頭蓋骨と顔の構造を持っていた。第3指はより強靭になり体重を支えていた。パラヒップスの歯冠は鋭くなった。
メリキップス


中新世中期にはメリキップス(Merychippus)が繁栄していた。メリキップスにはステップ地帯の硬い草をすり潰すのに使用されたとされるパラヒップスより大きな大臼歯があった。後脚の第2指と第4指は短くなり、走るときだけ地面に触れていたと考えられている。メリキップスは少なくとも19種類の種に分岐した。
ヒッパリオン

メリキップスから分岐したウマ科の中で特に変化があったのが、ヒッパリオン(Hipparion)、プロトヒップス(Protohippus)、プリオヒップス(Pliohippus)の3種である。メリキップスからもっとも変化したのがヒッパリオンで、歯冠のエナメル質が舌を隔離する壁を作っていた。北米で見つかったヒッパリオンの完全な化石は小さいポニーほどのサイズだった。化石から、カモシカのように体重が軽く乾いた大草原での生活に適応していたことがわかっている。脚の指は3本あったが、第3指のみで歩行し、第2指、第4指はすでに使われていなかった。
北アメリカではヒッパリオンとその近縁種(コーモヒッパリオン(Cormohipparion)、ナニップス(Nannippus)、ネオヒッパリオン(Neohipparion)、スードヒッパリオン(Pseudhipparion))は繁栄した。また、中新世の間に、アジアやヨーロッパにわたっている(ヨーロッパではアメリカの化石よりも小さなヒッパリオンが見つかっている。特にアテネで見つかった化石が有名である)。
シマウマやロバなどにはヒッパリオンから進化したという説が有力である。
プリオヒップス

プリオヒップス(Pliohippus)は約1200万年前、中新世中期にカリップス(Calippus)から進化した。プリオヒップスはエクウスとよく似た外見をしていた。蹄の両側にある2本の指は、ほぼ完全に退化しており、脚としての機能は完全に消失していた。プリオヒップスは長く細い脚を手に入れたことで、より速く走ることができるようになった。
プリオヒップスは解剖学的に多くの同一性が見られることから、現代の馬の先祖であると考えられてきた。しかし、プリオヒップスの頭骨には深い窪みがあるが、エクウスには窪みは見られず、現代の馬がまっすぐな歯であるのに対し、プリオヒップスの歯は曲がっていた。このような相違点が見られることから、プリオヒップスは現代の馬の先祖でない可能性もある。プリオヒップスはアストロヒップス(Astrohippus)の先祖である可能性がある。
ディノヒップス
ディノヒップス(Dinohippus)は鮮新世後期の北米で最も一般的なウマだった。元々、蹄は一つであると見られていたが、1981年にネブラスカで見つかった化石より、指は3本あったことが確認されている。
プレシップス
プレシップス(Plesippus)はディノヒップスとエクウスの中間種であると考えられている。
アイダホ州ヘイガーマンの近くで見つかった化石は、ヘイガーマン化石層群の約350万年前、鮮新世の地層から出土し、元々はプレシップスの近縁種のものと考えられていた。化石はプレシップス模式種(Plesippus shoshonensis)と呼ばれていたが、古生物学者らによる研究で化石はエクウスに最も近い化石であることが確認された。化石から算出した平均体重は425キログラムで、アラブ種と同じサイズだった。
鮮新世後期に北米の気候が変化し、北米で生活していた動物のほとんどは移動した。プレシップスも例外ではなく、約250万年前に陸続きだったベーリング地峡を渡り、ユーラシア大陸へと渡った。
エクウス
エクウス(Equus)の最古の種はエクウス・ステノニス(Equus Stenonis)で、イタリアで発見された。第四期の初期か新第三期の最後にプレシップスから進化したと考えられている。エクウス・ステノニスは後に体重が軽い種と重い種の2種に分岐した。
エクウス・ステノニスは北米に渡った。北米ではエクウス・スコッティ(Equus scotti)と呼ばれており、現代の馬を超える大きさの化石も見つかっている(エクウス・スコッティ・ギガンテウス(Equus scotti var.giganteus)と呼ばれる)。

アメリカ大陸では鮮新世(300万年前)から更新世(1万年前)までの間で、ヒッピディオン(Hippidion)、エクウス・フランシスキ(Equus francisci)、そして現代の馬の先祖の、大きく分けて3種類に分岐した。
ヒッピディオンは約300万年前にアメリカ大陸間大交差が起こった際、北米を渡り南米で繁栄した。ヒッピディオンは比較的短い脚を持っていた。元々はプリオヒップスから進化したと考えられていたが、DNAの研究が進みエクウスに属する種であることが示されている。100万年前、南米にエクウスカバルス(Equus caballus)が渡ってくると、ヒッピディオンとエクウスカバルスは血が混じりあい、一つの種となった。ヒッピディオンは南米の固有種となっていたが、約1万3000年前に絶滅した。
エクウスフランシスキは更新世の氷床の南で見られた。エクウスフランシスキは約3万1000年前、ベーリング地峡周辺で絶滅した。
Equus ferus

エクウスはドイツやシベリアで生活していた現代の馬に連なるウマ(ノウマ、Equus ferus)と、アラスカで生活した全北区の種の2種に分かれた。
現代の馬に連なるウマ(ノウマ、Equus ferus)は中央ヨーロッパから北米まで広く繁栄し、現代の馬へと進化していった。なお現生の家畜馬の染色体数は64本だが、モウコノウマ(ノウマの一亜種)については染色体数は66本である。
全北区の種は北米から離れることなく生活していたが、人類がウマを家畜とする以前に絶滅したと見られている。
西カナダに、北米の全てのウマ科が1万2000年前から1万1000年前にかけて絶滅したという痕跡が残っている。このときは、アメリカで生活していたほとんどの大型生物も同時期に絶滅しており、しばしば議論の的となる。大量絶滅とそれまでの哺乳類の繁栄を比べると、その衰退は激しく、その要因として主に2つの仮説が考えられている。1つは、気候変動により約1万2500年前にステップ地帯の植物がツンドラによって枯れ食料がなくなり絶滅したという仮説と、もう1つは馬が絶滅した時期と、クローヴィス文化の発祥がほぼ同時である事から人類によって狩られたという仮説である。
馬の化石は1万年前の地層以降しばらく発見されていなかったが、カザフスタンとウクライナ南の6000年前の地層から再び出土し始めた。6000年前から、馬を飼育したり、騎乗し始める人々がでてきた可能性が高い。その方法は比較的早く広まり、いくつかの部族では馬がその生活において重要な役割をしていた可能性がある。
なお、北アメリカにはマスタングと呼ばれる野生馬があるが、これはすべて1493年にクリストファー・コロンブスが再導入し、その後複数回持ち込まれたものを起源として、それらが野生化したものである。アメリカ大陸の原住民には、馬を指し示す言葉がなかったため、犬や鹿の一種として呼ぶようになった(たとえばヘラジカ犬(elk-dog)など)。

特徴

歯
馬の進化の特徴として、歯が常に変化していたことが挙げられる。雑食性を示す短くでこぼこな大臼歯から、草食性哺乳類に共通な長く平坦な歯へと徐々に変化していった。そのほかの変化に、頭骨の顔の部分が伸びたこと、短かった首は脚が長くなるにつれ長く伸びたことが挙げられる。それらの要因によって、最終的に馬の体のサイズは大きくなった。
指
馬の先祖は第3指とその隣の指を使って歩いていた。その骨格は腓骨とよばれる中手骨、中足骨の側面に痕跡として残っている。基本的には意味を成さないが、手根関節と足根骨の飛節を支える重要な役割を果たしているという意見もある。
知能
関連項目
- 三趾馬
 ウィキニュースに関連記事があります。ドイツで発掘されたウマ科の化石「Eurohippus messelensis」に関する論文が投稿、子宮が残存した化石としては世界最古(2015年11月16日)
ウィキニュースに関連記事があります。ドイツで発掘されたウマ科の化石「Eurohippus messelensis」に関する論文が投稿、子宮が残存した化石としては世界最古(2015年11月16日)
脚注
This article uses material from the Wikipedia 日本語 article ウマの進化, which is released under the Creative Commons Attribution-ShareAlike 3.0 license ("CC BY-SA 3.0"); additional terms may apply (view authors). コンテンツは、特に記載されていない限り、CC BY-SA 4.0のもとで利用可能です。 Images, videos and audio are available under their respective licenses.
®Wikipedia is a registered trademark of the Wiki Foundation, Inc. Wiki 日本語 (DUHOCTRUNGQUOC.VN) is an independent company and has no affiliation with Wiki Foundation.