Nucleocytoviricota: Gruppe von Viren
Das Phylum der Nucleocytoviricota (früher auch englisch Nucleocytoplasmic large DNA viruses, NCLDV) umfasst eine heterogene Gruppe meist großer dsDNA-Viren, die eine Reihe bestimmter Gene (NCLDV core genes) aufweisen, die gewöhnlichen Viren fehlen.
Gründungsmitglieder der Gruppe (2001) sind die Familien Asfarviridae, Iridoviridae und Poxviridae (Pockenviren), sowie die Phycodnaviridae. Weitere Mitglieder sind die Familien Mimiviridae (synonym mit „Megaviridae“), Marseilleviridae, Ascoviridae. Vorgeschlagene Mitglieder sind die Pithoviren, Orpheoviren und Pandoraviren, gegebenenfalls mit jeweils eigene Familien, sowie etliche mehr. Mögliche Kandidaten sind Dinodnavirus und „Urceolovirus“. Diese Mitglieder bilden – wie inzwischen mehrfach bestätigt – eine monophyletische Verwandtschaftsgruppe, d. h. sie haben einen gemeinsamen viralen Vorfahren. Man hatte daher zunächst verschiedentlich vorgeschlagen, diese Gruppe als „Megavirales“ in den Rang einer neuen Virusordnung zu erheben. Nachdem das International Committee on Taxonomy of Viruses (ICTV) seit 2018 auch Ränge oberhalb dem der Ordnung zuließ, wurden aufgrund der hohen Diversität dieser Gruppe solche höheren Ränge favorisiert. Damit wurde es ermöglicht, die Hauptgruppen der NCLDV als Ordnungen oder gar Klassen aufzustellen. So schlugen beispielsweise Guglielmini et al. (2019) die erweiterte Familie Mimiviridae auf als eine Ordnung und schlugen dafür die nun frei gewordene Bezeichnung „Megavirales“ vor. Diese Entwicklung erreichte 2019/2020 mit der Master Species List Nummer 35 des ICTV ihren vorläufigen Abschluss mit der offiziellen Anerkennung der NCLDV als Phylum Nucleocytoviricota (ursprünglich „Nucleocytoplasmaviricota“) mit den Klassen Megaviricetes und Pokkesviricetes durch das ICTV im März 2020. Die Ordnung der erweiterten Mimiviridae heißt jetzt Imitervirales.
| Nucleocytoviricota | ||||||||
|---|---|---|---|---|---|---|---|---|
 EM-Aufnahme eines Virions (Virusteilchens) des | ||||||||
| Systematik | ||||||||
| ||||||||
| Taxonomische Merkmale | ||||||||
| ||||||||
| Wissenschaftlicher Name | ||||||||
| Nucleocytoviricota | ||||||||
| Kurzbezeichnung | ||||||||
| NCLDV | ||||||||
| Links | ||||||||
|
Genom
Das Genom der NCLDV ist vielfältig und hat eine Größe im Bereich 100 bis über 1500 kb (Tupanvirus). Viele NCLDV sind damit Riesenviren (englisch giant viruses, giruses), wofür meist eine Überschreitung der 300 kb-Grenze vorausgesetzt wird (bei Yutin und Koonin von 500 kb). Umgekehrt sind alle Riesenviren (bisher) dsDNA-Viren im Phylum NCLDV. Die größten Vertreter bilden größere Viruspartikel und haben ein größeres Genom als viele kleine Bakterien – die Mimiviren wurden zunächst sogar für (parasitäre) Bakterien gehalten, zumal sie auch Gram-Färbung zeigen. Zur späten Entdeckung der meisten Riesenviren (zahlreich erst ab etwa 2003) trug bei, dass sie bei der Suche nach Viren in den Filtern (mit typischer Porengröße von 0,2 μm) hängen blieben, die Bakterien und Protisten von Viren abtrennen sollten, langsamer zu leicht sichtbaren Klumpen aggregieren und sich auch langsamer vermehren. 2020 berichteten Wissenschaftler Stoffwechselgene in den metabolismuslosen Viren gefunden zu haben, was darauf hindeutet, dass diese den Metabolismus ihrer Wirtszellen ändern.
Wirte
Obwohl die NCLDV mehr eigene Proteine kodieren als gewöhnliche Viren (oft Hunderte statt kaum ein Dutzend), sind sie wirtspezifisch. Unter den Wirten sind ausschließlich komplex-zelluläre Organismen (Eukaryoten): viele Protisten (z. B. Amöben und Algen), aber auch Wirbeltiere und Insekten. Ursprünglich sechs, nun neun gemeinsame (homologe) Gene sind in allen NCLDV zu finden (NCLDV core genes), 177 weitere Gene (Stand 2009) kommen in mindestens zwei der Familien vor. Dazu gehören vier Gene, welche die DNA-Replikation und Reparatur-Mechanismen betreffen: die DNA-Polymerase-Familie B, die Topoisomerase II A, die „Flap“-Endonuklease und das Ringklemmenprotein (Proliferating-Cell-Nuclear-Antigen) sowie die RNA-Polymerase II und den Transkriptionsfaktor II B. Die Gene mancher NCLDV enthalten auch Introns.
Vermehrungszyklus
Manche der NCLDV vermehren sich ganz oder teilweise im Zellplasma (Zytoplasma) der eukaryotischen Wirtszellen, andere gehen möglicherweise durch eine nukleare Phase im Zellkern. Viele NCLDV bilden nach der Infektion im Zytoplasma ihres Wirtes eine mikroskopisch sichtbare Produktionsstätte (Virusfabrik, englisch virus factory) aus. Bei diesen gibt es zum Teil andere Viren, die deren Syntheseapparat für ihre eigene Vermehrung nutzen und daher Virophagen (‚Virenfresser‘) genannt werden. Der erste entdeckte Fall dieser Art war Sputnikvirus 1 mit dem NCLDV Acanthamoeba castellanii mamavirus (AcMV), weitere Virophagen wie ‚Zamilon‘ befallen ebenfalls Vertreter der Mimiviridae. Zwar werden Viren traditionell und meist auch heute noch nicht als Lebewesen angesehen, die durch NCLDV vollbrachten Leistungen verringern aber die Kluft zur belebten Welt. Unverändert gültig ist, dass Virionen keinen Stoffwechsel aufweisen, der auf ATP als „Energiewährung“ beruht.
Horizontaler Gentransfer
Zwischen den Riesenviren als viralen Endocytobionten (Organismen, die in den Zellen anderer Organismen leben oder sich vermehren) und ihren oft amöboiden Wirten lässt sich massiver horizontaler Gentransfer in beiden Richtungen (AtoV, Amöbe auf Virus und umgekehrt VtoA, Virus auf Amöbe) nachweisen.
Die Tatsache, dass für einige der unter den NCLDV verbreiteten Genen keine Entsprechung in zellulären Organismen gefunden wurde, wurde vom Team um Didier Raoult als Hinweis auf eine vierte Domäne des Lebens gedeutet, deren Vertreter bis auf die parasitierenden NCLDV ausgestorben sei. Diese Forth-Branch-of-Life-Hypothese ist nicht mehr haltbar, seit für mehrere in NCLDV-Gruppen verbreitete Gene Abstammungen von weit auseinander liegenden Stellen im Baum des Lebens gezeigt wurden, überwiegend von verschiedenen Eukaryonten, aber auch von Bakterien. Allerdings beflügelt die Entdeckung jeder neuer Familie der Riesenviren die Diskussion von Neuem, wie das Beispiel der Gattung Medusavirus zeigt.
Eine Zusammenfassung dieser Diskussion findet sich bei Traci Watson (2019).
Es wurde in diesem Zusammenhang sogar nachgewiesen, dass es nicht nur einen Gentransfer zwischen den amöboiden Wirten und den Riesenviren als viralen Endocytobionten gibt (VtoA/AtoV), sondern sogar zwischen den Viren und gleichzeitig vorhandenen bakteriellen Endocytobionten. Möglicherweise könnte dies die merkwürdige Übereinstimmung im Abwehrsystem CRISPR der Bakterien gegenüber Viren (also Bakteriophagen) einerseits und dem Abwehrsystem MIMIVIRE der Mimiviren gegenüber parasitierenden Satellitenviren (also Virophage) andererseits erklären.
Endogenisierung
Die Landpflanzen Physcomitrella patens (Laubmoose) und der Selaginella moellendorffii (Lycophyten) besitzen offene Leserahmen (open reading frames, ORFs), die von endogenisierten NCLDVs einer noch unbekannten Familie stammen könnten.
Histone
Eine Reihe von NCLDVs kodiert Histone oder Histon-Homologe. Beispiele finden sich in den Familien Marseilleviridae, Phycodnaviridae mit „Pandoravirus“ und „Clandestinovirus“, Iridoviridae, Mamonoviridae und Unterfamilie Klosneuvirinae. Anderweitige Beispiele gibt es in der Gattung Bracovirus (alias Bracoviriform, Spezies „Cotesia vestalis bracovirus“, CvBV, zu NCLDV?) und der Familie Nudiviridae.2022 wurde die Rolle von Histonen im Replikationszyklus einer Reihe von Viren umfassend untersucht.
Systematik
Innere Systematik und Forschungsgeschichte
Trotz ihrer Diversität bilden die NCLDV wie mehrfach bestätigt eine Verwandtschaftsgruppe (Taxon), wobei die Riesenviren von (verschiedenen) Gruppen kleinerer NCLDV abzustammen scheinen, statt umgekehrt.
- „Irido-Ascoviridae“: Mehrere Studien unterstützen seit dem Jahr 2000 die Annahme, dass die Ascoviridae sich aus den Iridoviridae entwickelt haben, es könnte aber auch umgekehrt sein. Weiter wird vermutet, dass sich aus den Ascoviridae die Gattung Ichnovirus (Familie Polydnaviridae) entwickelt hat.
- Dinodnavirus: Untersuchungen des Genoms von Heterocapsa circularisquama DNA virus 01 (Gattung Dinodnavirus) haben 2009 gezeigt, dass diese ursprünglich in die Familie Phycodnaviridae gestellte Gattung eher zur Familie der Asfarviridae gehört.
- „Urceolovirus“: Eine weitere Spezies von DNA-Riesenviren, „Urceolovirus corneum“ („KLaHel“), erstmals berichtet 2015, könnte Mitglied der NCLDV sein.
- Klosneuvirinae: Im April 2017 wurde über den Fund von vier neuen Riesenviren in der Kläranlage Klosterneuburg berichtet. Michael Wagner (Department für Mikrobiologie und Ökosystemforschung der Universität Wien), Holger Daims, Matthias Horn und Frederik Schulz (alle Uni Wien) publizierten über diese inzwischen vom ICTV bestätigte Unterfamilie Klosneuvirinae der Mimiviridae in Science.
- „Orpheovirus“: Im Januar 2018 wurde von Julien Andreani et al. über ein neues Riesenvirus Orpheovirus berichtet. Nach Vorschlag der Autoren soll „Orpheovirus“ in eine eigene Familie „Orpheoviridae“ gestellt werden, in enger Verwandtschaft zur vorgeschlagenen Familie Pithoviridae der Pithoviren.


- „Meelsvirus“: Im September 2018 fanden Shinn und Bullard bei der nochmaligen Analyse von elektronenmikroskopischen Aufnahmen aus den 1980er Jahren ein Riesenvirus („Meelsvirus“), das den Pfeilwurm Adhesisagitta hispida infiziert, im Zellkern repliziert und dessen Virionen bei einer Länge von 1.25 μm umhüllt sind. Wegen fehlender Genomdaten ist eine bessere Einordnung bislang nicht möglich.
- Waldbodenviren: Im November 2018 berichteten Frederik Schulz und Kollegen über die Entdeckung von 16 neuen Riesenviren per Metagenomanalyse von Waldbodenproben, die sich nur teilweise bekannten Gruppen zuordnen zu lassen scheinen. Für diese wurden vorläufige Namen vergeben, die meist auf ihre Herkunft hinweisen, unter anderem „Faunusvirus sp.“ (nicht zu verwechseln mit der offiziellen Gattung Faunusvirus der Familie Chaseviridae), „Gaeavirus“, „Homavirus“, „Barrevirus“, ‚Edafosvirus‘, „Hyperionvirus“, „Harvfovirus“, „Terrestrivirus“, „Dasosvirus“, „Satyrvirus“ (alle Mimiviridae), sowie „Sylvanvirus“, „Solivirus“ und „Solumvirus“. Möglicherweise ist dies nur die „Spitze eines Eisbergs“.
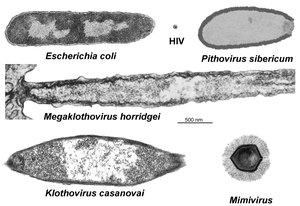
- Yasminevirus etc.: Im März/April 2019 berichteten Clara Rolland und Kollegen in einer Zusammenfassung von Forschungsergebnissen über Riesenviren erstmals über weitere Kandidaten: „Clandestinovirus“, Fadolivirus, „Sissivirus“, „Usurpativirus“, Yasminevirus und „Misannotatedvirus“ (informell auch „misidentified virus“); einem „mine drainage virus“; sowie dem Virophagen Sissivirophage. Die Gattungen Fadolivirus und Yasminevirus wurden im April 2023 vom ICTV bestätigt.
- „Choanovirus“: Im September 2019 berichteten David M. Needhal, Alexandra Z. Worden et al. über zwei Spezies (1 und 2) einer weiteren neuen Gattung „Choanovirus“ der erweiterten Mimiviren (jetzt Imitervirales). Der nächste Verwandte könnte das Aureococcus anophagefferens virus (AaV, Kratosvirus quantuckense) sein.
- „Klothoviridae“: Ebenfalls 2019 schlugen Roxane-Marie Barthélémy et al. eine Familie „Klothoviridae“ mit Typusspezies „Klothovirus casanovai“ und einer weiteren Spezies „Megaklothovirus horridgei“ vor, die mit 2,5–3,1 μm und 4 μm einen neuen Größenrekord darstellen würden. Als Wirte dienen wie bei „Meelsvirus“ Pfeilwürmer. Ähnlich wie bei den Arenaviridae fanden sich Ribosomen in den Viruspartikeln. Da derzeit noch keine Genomdaten vorliegen ist eine genauere Einordnung der Familie noch nicht möglich.
.large.TMEs.jpg)
(D–F) Die Aufnahmen zeigen vergrößerte Zellkerne mit sich entwickelnden Viroplasmen.
(G–I) Die Morphologien der drei Viren umfassen einen Genom-Kern, umgeben von einem ikosaedrischen Nukleokapsid.

- „Mininucleoviridae“: Im Januar 2020 wurde von Subramaniam et al. eine weitere Familie „Mininucleoviridae“ von Riesenviren der NCLDV vorgeschlagen, deren Mitglieder Krebstiere (Crustacea) parasitieren. Zu den Mitgliedern dieser Familie gehören nach Vorschlag „Carcinus maenas virus 1“ (CmV1), „Dikerogammarus haemobaphes virus 1“ (DhV1) und „Panulirus argus virus 1“ (PaV1). Die Familie gehört offenbar zum Pitho-Irido-Marseille-Zweig der NCLDV (jetzt Ordnung Pimascovirales).
- Im Oktober 2020 wurde eine Metagenomanalyse veröffentlicht, die zeigte, dass die Korallenbleiche bei Mo'orea, Franz. Polynesien, im Zusammenhang mit Viren steht. Obwohl eine eindeutige Zuordnung des rekonstruierten „assembled coral giant virus“ zu einem bestimmten Vertreter der NCLDV nicht möglich war, steht eine Zugehörigkeit zu dieser Gruppe außer Frage. Weitere Untersuchungen sind nötig.
Systematik nach ICTV 2020

• Mimiviridae ist im erweiterten Sinn zu verstehen (Ordnung Imitervirales),
• die Asfar- und Poxviridae bilden eine Klade (Klasse Pokkesviricetes),
• die Ascovirdae sind paraphyletisch und bilden mit den Iridoviridae eine Klade („Irido-Ascoviridae“)
.svg)
Das Original wurde um die neuen ICTV-Bezeichnungen und den vorgeschlagenen Familiennamen ergänzt.

Das ICTV ist im März 2020 dem Vorschlag von Eugene Koonin et al. (2015 und 2019) einer inneren Systematik der NCLDV gefolgt, in der eine erweiterte Familie der Asfarviridae (jetzt Ordnung Asfuvirales) eine Schwestergruppe der Poxviridae (mit Ordnung Chitovirales) bilden. Von Koonin et al. (2015 und 2019) stammt die damit verträgliche Gliederung in drei Hauptgruppen (oder Zweige, englisch branches) wie folgt:
- Phylum Nucleocytoviricota (Nucleocytoplasmic large DNA viruses (NCLDV), ursprünglicher Vorschlag „Nucleocytoplasmaviricota“, ersetzt ältere Vorschläge als Ordnung „Megavirales“ s. l.)
- Klasse Megaviricetes
- Zweig 1:
- Ordnung Algavirales: die Familie Phycodnaviridae mit den Gattungen Pandoravirus, „Mollivirus“ und „Usurpativirus“, aber ohne umgruppierte Vertreter wie die „OLPG“ (Organic Lake Phycodna group)
- Ordnung Imitervirales (veraltet „Megavirales“ s. s.): Familie Mimiviridae (alias Megaviridae), Mesomimiviridae („OLPG“), Allomimiviridae („Tetraselmisviren“) und Schizomimiviridae („Aureococcusviren“)
- Zweig 2: Ordnung Pimascovirales (Zusammenziehung aus Pitho, Irido, Marseille und Asco, früher auch MAPI-Superklade oder PMI-Gruppe genannt). Von oben nach unten immer basaler werdend gehören dazu:
- die Familien Iridoviridae und Ascoviridae (derzeit vom ICTV noch getrennt aufgestellt, per Vorschlag eine gemeinsame Familie „Asco-Iridoviridae“), ggf. mit Gattung Ichnovirus (bisher Polydnaviridae)
- Familie Marseilleviridae
- die vorgeschlagene Familie „Mininucleoviridae“
- die vorgeschlagene Familie „Pithoviridae“ (mit „Cedratvirus“, „Orpheoviridae“, „Solumvirus“, „Solivirus“, „Sissivirus“, „Misannotatedvirus“ (informell auch „misidentified virus“) und „mine drainage virus“)
- Zweig Medusaviren: Familie Mamonoviridae (ursprünglich als Medusaviridae vorgeschlagen): Gattung Medusavirus und „Clandestinovirus“
- Zweig 1:
- Klasse Pokkesviricetes
- Zweig 3:
- Ordnung Asfuvirales: (erweiterte) Familie Asfarviridae mit Asfivirus (Erreger der Afrikanischen Schweinepest), „Faustovirus“, „Pacmanvirus“, „Kaumoebavirus“, evtl. Dinodnavirus, etc.
- Ordnung Chitovirales: Familie Poxviridae (Pockenviren)
- Zweig 3:
- Klasse Megaviricetes
In dieses Schema wurden die Kandidaten nach Schulz et al. (2018), Rolland et al. (2019) und Subramaniam (2020) eingetragen, soweit sie nicht zu den erweiterten Mimiviridae (Ordnung Imitervirales) gehören. Die Kandidaten Dinodnavirus und „Urceolovirus“ sind vom ICTV noch nicht berücksichtigt, was dem Vorschlag von Koonin et al. vom April 2019 entspricht.
Guglielmini et al. (2019) hatten alternativ die Einteilung in zwei Superkladen wie folgent vorgeschlagen:
- MAPI-Superklade (Zweig 2 von oben: Pimascovirales)
- PAM-Superklade (entspricht in etwa Zweig 1 und 3 (nur Asfuvirales))
Die Poxviridae (Pockenviren) und Mamonoviridae (Medusaviren) sind in diesem Vorschlag unberücksichtigt.
Nach dem früheren Vorschlag von Schulz et al. (2018), Fig. 2, stehen die Poxviridae dagegen basal in den NCLDV, die (erweiterten) Asfarviridae wurden im Zweig der Marseilleviridae verortet. Im Gegensatz zu Koonin et al. (2015 und 2019) erschienen die Riesenviren hier noch wie bei Guglielmini et al. monophyletisch. Das ICTV ist 2020 jedoch den Ergebnissen von Koonin, Yutin, Bäckström, Ettema et al. gefolgt, nachdem sich Riesenviren innerhalb der NCLDV mehrmals aus einfachen Vorstufen entwickelt haben:
Nach Clara Rolland et al. (2019), Fig. 2 bilden die Imitervirales mit den Pokkesviricetes eine gemeinsame Klade, Schwestergruppen sind die Algavirales und Pimascovirales gleichauf. Die Megaviricetes scheinen – obgleich im März 2020 vom ICTV als Taxon offiziell bestätigt – keine Klade als Schwestergruppe der Pokkesviricetes zu bilden.
Nach Disa Bäckström et al. (2019), Fig. 1, scheinen – im Gegensatz zu Fig. 6 – die Pokkesviricetes ähnlich wie bei Schulz et al. (2018) keine gemeinsame Klade zu bilden, die Poxviridae (bzw. Chitovirales) stehen basal, erst danach zweigen die Asfarviridae (bzw. Asfuvirales) vom Rest der NCLDV ab.
Weitere Stammbäume basierend auf Neighbor-joining und auf Maximum likelihood findet man u. a. in den folgenden Arbeiten:
- William H. Wilson et al. (2017), Fig. 2
- Claire Bertelli et al. (2017), Fig. 4
- Disa Bäckström (2018), Fig. 2
- Disa Bäckström et al. (2019), Fig. 1 und 6
- Sailen Barik (2018), Fig. 1
Äußere Systematik
Die Organisation des Genoms und der DNA-Replikationsmechanismus legen eine phylogenetische Beziehung nahe zwischen den Rudiviren (Ordnung Ligamenvirales: Rudiviridae) und großen eukaryalen DNA-Viren (NCLDVs) wie dem Afrikanische Schweinepestvirus (African swine fever virus, Asfarviridae), Chloroviren (Chlorella virus, Phycodnaviridae) und Pockenviren (Orthopoxvirus, Poxviridae).
Koonin et al. (2015, 2019) vermuten den Ursprung der NCLDV in den Tectiviridae, ikosaedrischen schwanzlosen ssDNA-Bakteriophagen, nach ICTV ebenfalls im Reich Bamfordvirae – im Unterschied zu den Herpesvirales (Herpesviren), bei denen der Ursprung bei den geschwänzten Caudovirales vermutet wird. Die Entwicklung verlief nach diesem Vorschlag über oder vermittels von Polintoviren (Polintons, auch Mavericks genannt: große DNA-Transposons, die virale Proteine kodieren, aber auch häufig in eukaryotischen Genomen vorkommen). Auch die Entwicklung von Adenoviren (Adenoviridae) und Bidnaviren (Bidnaviridae) sowie von Virophagen (der Familie Lavidaviridae) wurde, so die Vermutung, durch die Polintons initiiert. Die im März 2020 vom ICTV getroffenen taxonomischen Einordnungen tragen dem Rechnung: sie ordnen
- die Herpesvirales und Caudovirales dem Reich Heunggongvirae im Bereich Duplodnaviria zu;
- die Tectiviridae, Adenoviridae (beide Klasse Tectiliviricetes) und Lavidaviridae (Klasse Maveriviricetes) dem Phylum Preplasmiviricota, einem Schwesterphylum der NCLDVs im Bereich Varidnaviria zu;
- die Yaraviridae mit der Gattung Yaravirus den Bamfordvirae ohne nähere Zuordnung (incertae sedis);
- lediglich die Bidnaviridae wurden per Vorschlag – ihrem Namen zum Trotz – dem Bereich Monodnaviria zugeordnet.
Literatur
- Natalya Yutin, Yuri I. Wolf, Eugene V. Koonin: Origin of giant viruses from smaller DNA viruses not from a fourth domain of cellular life. In: Virology. 2014, doi:10.1016/j.virol.2014.06.032, PMC 4325995 (freier Volltext).
- N. Yutin, P. Colson, D. Raoult, E. V. Koonin: Mimiviridae: clusters of orthologous genes, reconstruction of gene repertoire evolution and proposed expansion of the giant virus family. In: Virol J. 10, 4. April 2013, S. 106, PMID 23557328
Weblinks
- Dagmar Röhrlich: Im Reich der Riesenviren. Deutschlandfunk – Wissenschaft im Brennpunkt, 27. Dezember 2015.
Anmerkungen
Einzelnachweise
This article uses material from the Wikipedia Deutsch article Nucleocytoviricota, which is released under the Creative Commons Attribution-ShareAlike 3.0 license ("CC BY-SA 3.0"); additional terms may apply (view authors). Abrufstatistik · Autoren Der Inhalt ist verfügbar unter CC BY-SA 4.0, sofern nicht anders angegeben. Images, videos and audio are available under their respective licenses.
®Wikipedia is a registered trademark of the Wiki Foundation, Inc. Wiki Deutsch (DUHOCTRUNGQUOC.VN) is an independent company and has no affiliation with Wiki Foundation.